Farmers and crop consultants are encouraged to complete this survey
This survey is intended to estimate the impact of pests on soybeans and currently used management strategies. Your perspective is valuable in making these estimates as accurate as possible. Your responses will be kept confidential and only aggregated responses for the state will be published. You can find last year’s estimates at https://midsouthentomologist.org.msstate.edu/Volume17/Vol-17-1_TOC.html.
Emily Zobel, Senior Agriculture Agent Associate | ezobel@umd.edu
University of Maryland Extension, Dorchester County
Corn
Continue scouting for cutworms and stink bugs. The threshold for cutworms is 2-5% cut plants up to the V5 stage and with active larvae present. Stinkbugs will move into corn as surrounding small grains are harvested. When scouting for stinkbugs, pay close attention to the thorax of brown color stink bugs as the beneficial spined soldier bug is often mistaken for the invasive brown marmorated stink bug. The spined soldier bug has a prominent spine on each “shoulder.” NCSU’s suggested threshold is 13 stinkbugs per 100 plants for V1-V6, 10 stinkbugs per 100 plants for V14-VT, and 28 stinkbugs per 100 plants for R1-R2. The most critical time to treat if between V14 to VT is just before the primary ear is exposed to avoid banana ear. Stinkbugs will likely be around the primary ear at this point so that should be the target of the application.
Soybean
Early-season defoliator pests that are active right now include bean leaf beetles, green cloverworms, slugs, and grasshoppers. Soybeans can typically withstand a decent amount of defoliation before yield losses occur. If defoliation reaches 30%, and you are finding one grasshopper per sweep or 2-3 bean leaf beetles per plant treatment may be advisable.
Figure 1. Bean leaf beetle with feeding damage on a soybean leaf. Photo by Adam Sisson, Iowa State University, Bugwood.org.
Alfalfa
Begin scouting for potato leafhopper. Ten sets of 10-20 sweeps using a sweep net should be taken in random locations in the field. A detail threshold for alfalfa is based on the plant size and cost of the hay can be found online but a general guideline is 3” or less is 20 leafhoppers per 100 sweeps, 4-6” tall is 50 per 100 sweeps, 7-10” tall is 100 per 100 sweeps, and greater than 11” is 150 per 100 sweeps. If the field is more than 60 percent bud stage or if it has experienced “hopper burn,” the alfalfa should be cut instead of sprayed. Fresh-cut alfalfa should not be sprayed as leafhopper adults tend to move out of the field when it is being cut.
Figure 2. Potato leafhoppers in a sweep net. Photo by Bryan Jensen, University of Wisconsin, Bugwood.org.
Maria Cramer, Galen Dively, and Kelly Hamby University of Maryland, Department of Entomology
It is not unusual to see groups of Japanese beetles feeding on corn silks, which is known as “silk clipping” Figs.1 and 2). While Japanese beetle numbers tend to peak in July, there are multiple beetles that may clip corn silks, and with later maturity field and sweet corn silking in August, it is important to still be on the lookout But how much of a concern is silk clipping, what should you be looking for, and what should you do about it?
Figure 1 (left). Japanese beetles feeding on corn silks. Figure 2 (right). Silk regrowth after clipping. Images: M. Cramer, University of Maryland
Silk clipping is often not as much of a concern as it initially appears. If silks are clipped after pollination, which occurs within the first 4-5 days of silk emergence1, kernel set will not be affected2. If clipping reduces the number of kernels, the kernels may develop to be larger and offset the reduction in number2. However, under drought conditions, yield loss from silk clipping is more likely2,3.
Drought slows silk emergence and pollination, which means there is a longer window where silk clipping can hurt yield. Indeed, severe drought stress can cause incomplete silk emergence and cause a mismatch between pollen shed and silks that results in nearly blank cobs1. Drought can also make it harder for plants to compensate for poor pollination1. If leaf rolling begins in the early morning and continues until evening1, the field is stressed enough to be of concern and it is important to scout for silk clipping beetles during the first several days of silk emergence.
The culprits. Japanese beetles are the most noticeable silk clippers in Maryland because they are large, shiny, and congregate in groups (Fig. 3). They are a sporadic pest4 and their populations will vary yearly. However, their populations may be higher in corn following sod, soybean, or perennial ryegrass or clover covercrops4. Other beetles that may clip silk include the western, northern, and southern corn rootworm adults (Fig. 4)5. Western corn rootworm (WCR) has several look-alikes that do not clip silks, so make sure check the stripes; WCR will not have crisp black stripes, but instead has smudged stripes.
Figure 3 (left). Japanese beetle. Image: E. Hodgson, Iowa State. Figure 4 (right). Adults of southern corn rootworm (left), western corn rootworm (middle), and northern corn rootworm (right). Image: Varenhorst, South Dakota State
Scouting. Silking typically begins 3 days after tasseling5, so plan your scouting accordingly. You want to evaluate the silk stage and pollination. Silks naturally senesce about 10 days after emergence, browning and drying out. At this point, pollination can no longer occur1. To determine if green silks have been successfully pollinated, you can dissect the ear and do a shake test. Pollinated silk starts to discolor and drop away at the base of the silk where it attaches to the ear. Bob Nielson with Purdue Extension has produced a great video describing the pollination shake test: https://www.youtube.com/watch?v=K7DiwD4N0T0&feature=youtu.be
You should scout if pollination is incomplete. When scouting, make sure you sample both the edges and the interior (at least 40 feet into the field); while you may see alarming numbers of Japanese beetles on the edge of the field, there are usually much fewer inside the field2. Sample a minimum of 20 corn plants in 5 locations spaced evenly though the field. Count the number of beetles per ear and measure the length of the silks.
Thresholds. For Japanese beetles, three conditions need to be met to before an insecticide application will pay off: 1) there are three or more beetles per ear, 2) silks are clipped to less than ½ inch in length, 3) and pollination is less than 50% complete4 (most silks in the field are still green and/or shake test indicates about half of the silks are still attached). Conditions are similar for rootworm beetles, but the threshold is five or more beetles per ear.
Treatments. Because broad-spectrum insecticides may cause flare ups of other pests (for example, aphids or spider mites), only spray if thresholds are met. Pollen-shed is a time when there are large numbers of beneficials in the corn field doing important pest control work (Fig. 5), and foliar sprays may decrease their numbers.
Figure 5. Lady beetle larva eating corn pollen. Image: M. Cramer, University of Maryland
For Japanese beetles, consider a perimeter spray if most of the damage is on field edges (where they tend to feed more heavily). Japanese beetles are difficult to control, but pyrethroids should provide some control (e.g., Baythroid®, Brigade®, Warrior II®, Hero®, etc.). Good adult corn rootworm control has been found for indoxacarb products (e.g., Steward®), pyrethroids (e.g., Warrior II®, Brigade, etc.), and neonicotinoid pyrethroid mixes (e.g., Endigo®)6,7. When using insecticides, always consult and follow the label.
If silk clipping by Japanese beetles is a consistent problem, consider cultural controls like avoiding ryegrass and clover cover crops. Because female beetles lay eggs more easily into soft ground, it is also possible to reduce egg laying in nearby fields by pausing irrigation during the peak of Japanese beetle activity4.
Steckel, S., Stewart, S. D. & Tindall, K. V. Effects of japanese beetle (Coleoptera: Scarabaeidae) and silk clipping in field corn. J. Econ. Entomol.106, 2048–2054 (2013). https://academic.oup.com/jee/article/106/5/2048/878220
Shanovich, H. N., Dean, A. N., Koch, R. L. & Hodgson, E. W. Biology and Management of Japanese Beetle (Coleoptera: Scarabaeidae) in Corn and Soybean. J. Integr. Pest Manag.10, (2019). https://academic.oup.com/jipm/article/10/1/9/5454734
DeVries, T. A. & Wright, R. J. Evaluation of Foliar Applied Insecticides for Control of Adult Corn Rootworm in Corn, 2015: Table 1. Arthropod Manag. Tests41, tsw080 (2016). https://academic.oup.com/amt/article/41/1/tsw080/2658080
DeVries, T. A. & Wright, R. J. Evaluation of Foliar-Applied Insecticides for Control of Adult Corn Rootworm in Corn, 2015C: Table 1. Arthropod Manag. Tests41, tsw096 (2016). https://academic.oup.com/amt/article/41/1/tsw096/2658095
Kelly Hamby1, Maria Cramer1, Galen Dively1, Sarah Hirsh2, Andrew Kness2 Alan Leslie2, Kelly Nichols2, Emily Zobel2, and David Owens3 1University of Maryland, Department of Entomology | 2University of Maryland Extension 3University of Delaware Extension
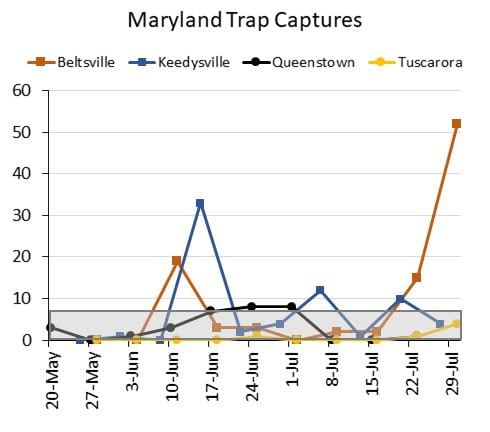
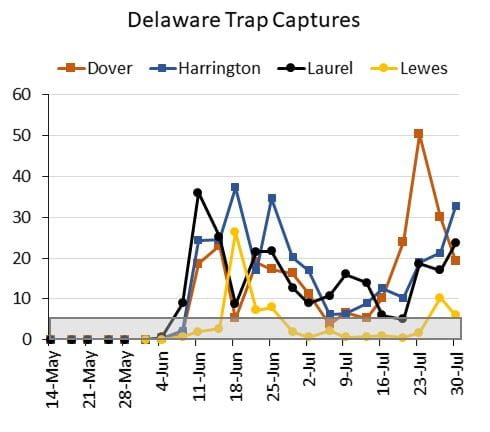
A few hot spots where corn earworm (also known as tomato fruitworm, soybean podworm, and sorghum headworm) activity is starting to rise have been identified in central Maryland and Delaware. The warm 2019-2020 winter allowed for overwintering in our area, and some parts of the state experienced a higher than normal first flight in early June. The warm weather through June and July made for speedy development and earlier activity for the second summer generation. Because corn earworm has developed resistance to most Bt hybrids, significantly more adult moths are emerging compared to levels a decade ago. Some areas continue to capture few moths and are experiencing low pressure, while others have been experiencing moderate pressure that may continue to increase towards heavy pressure (>65 moths captured per 5 days). Captures for select sites in Maryland and Delaware are pictured below, and values within the gray box indicate low pressure (<7 for weekly captures, and <5 for four to five day captures).
Corn earworm larva feeding damage to corn
Although corn earworm prefer fresh corn silks for egg laying, they will lay eggs on wilted and brown silks if the plants remain green and unstressed. As corn matures further over the next several weeks, corn earworm activity will shift to other host plants including soybeans and vegetables. See last summer’s articles for scouting and management recommendations in vegetables as well as sorghum and soybeans.
Podworm outbreaks have historically occurred in growing seasons where the corn crop was drought and heat stressed, with corn senescing earlier than normal. However, narrow row spacing in soybeans makes the plants less attractive to female moths and increases the likelihood that fungal pathogens will infect the larvae. Therefore, it is important to scout bean fields, especially paying attention to those fields with a more open canopy in areas where the nearby maturing corn is no longer attractive to earworm moths. North Carolina State University has produced a helpful economic threshold calculator for podworm in soybean: https://www.ces.ncsu.edu/wp-content/uploads/2017/08/CEW-calculator-v0.006.html
Acknowledgements: Corn earworm trapping efforts in were supported by the Crop Protection and Pest Management Program [grant numbers 2017-70006-27171 and 2017-70006-27286] from the USDA National Institute of Food and Agriculture. Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the view of the U.S. Department of Agriculture.
Emily Zobel, Agriculture Agent Associate University of Maryland Extension, Dorchester County
Soybean: The usual defoliators are starting to arrive, including bean leaf beetle, Japanese beetle, grasshoppers, and caterpillars. Control may be needed if there is 30% defoliation during the seedling and vegetative stages and 15% defoliation once plants start to bloom through pod fill.
Adult Dectes Stem borer will be emerging over the next several weeks. Chemical control is not recommended since it would require multiple applications to reduce larval infestations, which is not economical. If a high number of adults are found, harvesting that field as soon as it matures will reduce losses associated with lodged plants.
Fields that have an open canopy, drought-stressed, or have recently had an insecticide applied are at higher risk for corn earworm (CEW). CEW larva can feed on flowers without impacting yields because soybeans overproduce flowers. However, feeding during pod development can affect yield. An economic threshold calculator is available to assist with management decisions: https://soybeans.ces.ncsu.edu/wp-content/uploads/2017/08/CEW-calculator-v0.006.html.
Field Corn: As corn ears begin to form, check for stink bugs. Stink bugs will gather around the edges of fields, so scouting should be done at least 15 rows in. Thresholds are 1 stink bug per 4 plants when the ear is forming, and 1 stink bug per 2 plants from pollen shed to blister stage. Treatment is not recommended past the blister stage. Japanese beetles are minor defoliators and will clip corn silks, but control is not needed unless silks are cut back to less than ½ inch, and less than half the field has been pollinated.
Alfalfa: Once plants have hopper burn, there is no way to undo it, so continue scouting for leafhopper. Since infestations are highly variable, individual fields should be scouted. If you are planning on selling your hay for horse feed, check for blister beetle as well since they produce cantharidin, which causes skin blisters on humans and can make horses sick.
Sorghum: Sugarcane aphids were found on the Eastern Shore last year and typically show up in fields late July and August. Check underside of leaves for insects. Honeydew will turn leaves shiny and is an easy to see indicator that aphids are present. Sugarcane aphids are light yellow with black cornicles, antennae, and feet. Thresholds depend on plant growth stage; at boot to milk, thresholds are 50 aphids per leaf on 25 – 30% of plants. There is documented resistance to resistance to pyrethroids.
Cerruti R2 Hooks$ and Anahí Espíndola* $Associate Professor and Extension Specialist, CMNS, Department of Entomology *Assistant Professor, CMNS, Department of Entomology. Twitter: @Analyssi
Note: This is the third article of our series on pollinators. The initial articles can be found at the vegetable and fruit headlines news June special editions and Maryland Agronomy News Blog.
Introduction
Much has been written about bee (Hymenoptera) and butterfly (Lepidoptera) pollinators. Flies (Diptera) have also gotten their fair share of press regarding their pollination services. However, beetles (Coleoptera), which represent the largest insect order and are among the first flower-visiting insects in history, don’t receive similar admiration from pollinator enthusiasts and paparazzi. Nevertheless, even now, beetles are considered essential pollinators; and are especially important pollinators of some of the first flowering plants to evolve, such as magnolias and spicebush (Fig. 1). Beetles are attracted mostly to flowers that emit musky, yeasty, spicy, rotten or fermented odors. It just so happens that spicebush and magnolia flowers contain spicy pollen and produce aromatic oils, respectively, each of which serves as a lure for their beetle pollinators.
Fig. 1. Asian multicolored lady beetle on spicebush. Photo: J. Gallagher (CC)
There are 380,000 named living species of beetles, constituting nearly one-fourth of all known animal species on our planet. Beetles are so abundant that there are more species of beetles than of any other known group of animals. Among insects, beetles make up about 40% of all known species, and this may be due to their amazing ecological diversity, which allowed them to evolve impressive diverse morphological features. In the context of pollination, due to their sheer numbers, beetles contribute to a considerable amount of plant pollination. Even though the most consistent pollinators today belong to other insect orders, beetles do contribute significantly to the pollination of specific plants. Further, because beetles were one of the first insects to establish pollen-based relationships with plants, they have been described as key contributors to the reproduction of the first groups of flowering plant species.
Beetle history as pollinators
Beetles have been a predominant group of floral visitors and pollinators since very early times in the evolution of plants. Beetles have been evolving for roughly 250 million years, and are recognized among the first insects acting as pollinators of flowering plants. Indeed, until recently, many beetles and other insects were identified as pollinators of non-flowering plants (e.g. conifers, ginkgos, cycads). However, many ancient fossils indicate that plants today that are mostly wind-pollinated, relied strongly on insects for pollination in the past. Interestingly, a very recent study of fossil beetles found in amber (like the fossil insects from Jurassic Park!) provided the oldest account of flowering plant pollination. In this study, four different species of extinct beetles having lived 99 million years ago were found to carry pollen of flowering and non-flowering plants. While the non-flowering plant pollen corresponded to different species of cycads, pollen from flowering plants belonged to a newly described species of water lilies (Fig. 2). This not only pushed back the time of the first known fossil record of pollination, it also represents a clear link of how pollination of flowering plants may have evolved from pollinators that were visiting non-flowering plants. In comparison to other insect species currently known to visit flowers, beetles are now the oldest known pollinators of flowering plants, predating the oldest pollen-rich bee fossil by roughly 30 million years.
Fig. 2. Left: Water lily; Right: Cycads. Photos: A. Scottow and K. Norstog, respectively (CC).
Modern-day beetle pollinators visit many different types of plants, feeding on pollen, floral parts, and sometimes on nectar. While most beetles that act as pollinators are not specialized in their floral choices, beetles that visit flowers have some morphological traits that make them relatively efficient in transferring pollen. In fact, many of these beetles are hairy, allowing for pollen grains to stick on them and be transferred among plants. Further, some beetle groups have modified mandibles, with a brush-like structure or elongated proboscis, which allows them to more readily collect pollen and feed on nectar, respectively. While the vast majority of generalist beetle species visit well known plants such as carrot, buttercup, sunflower or cabbage, some beetle species are specialist pollinators of descendants of plants they visited millions of years ago: water lilies, custard apples, magnolias, all spice plants. For these latter plants, it is suspected that the evolution of beetles that they are associated with was driven by the diversification of their host plants. Relative to this point, part of the amazing morphological diversity that we see in extant beetle species is due to their long-dated interaction with these plants. Further, because these latter plant groups are usually associated with tropical or Mediterranean environments, it has been assumed for a long time that beetle pollination is most important in warm climates.
Beetles and magnolia’s special bond
Pollination by beetles seems to have strongly influenced the evolution of angiosperm flowers. Many million years ago, plants and pollinators began specializing and adapting traits that benefited each other’s requirements; and beetles were among the earliest pollinators to take part in nature’s matching adventures. Thus, flowers of lineages such as magnolias are paired with beetles as their primary pollinators (Fig. 3). Today, this flower’s characteristics are recognized as those that make up a “typical” beetle flower. These flowers are bowl-shaped and provide a cave-like structure for beetles to use as shelter. Beetles pollinating these flowers feed on pollen and sweet floral secretions. Though Magnolias from some areas are pollinated by several beetle families, some Magnolias from tropical areas have very specialized interactions, and are only effectively pollinated by few beetle species.
Figs. 3. Long horned beetle on Magnolia grandiflora. Photo: D. Hill (CC)
But how do beetles pollinate Magnolias? The story goes that beetles enter the flower carrying pollen from a previous floral visit, and passively deposit the pollen onto female structures while roaming about in the flower and feeding on stigmatic secretions. While these visits are happening, pollen of the flower is released, and while beetles feed on it, they also get it all over themselves. At the end of the flowering cycle, the flower wilts, and the messy beetles move on to another wide-open flower, where pollen is transferred and collection from the new flower initiates. This relationship between Magnolias and their beetle pollinators has evolved over millions of years of interactions. For example, magnolia plants have evolved strategies to “direct” beetle feeding away from reproductive parts of the flower. Basically, the plant sacrifices pollen and leaf tissue for the service of pollination. More specifically, beetles feed on other areas of the flower and are thus distracted from feeding on female parts of the flower which needs to stay in tacked so that it can produce seeds. Related to this, it has been suggested that this beetle-pollination relationship has led to an increase in the number of stamens produced by the plant, which insures that some are still present and ready to participate in plant reproduction after beetles finish their feast. Moreover, magnolia has developed securities that ban access by other insects during critical pollination phases and produces a fruity-smell that attracts beetles to the flowers.
Why do beetles visit flowers?
To understand beetle pollination, it is important to realize that most beetles visit flowers to feed on pollen, and sometimes on floral structures (e.g. petals, anthers) or secretions (e.g. stigmatic secretions). In fact, beetles rarely visit flowers for the typical nectar that other famous pollinators seek, and this reward is actually often absent or moderately produced in flowers beetles frequent. The most important reward beetles are after when visiting flowers is protein-rich pollen. Pollen beetles (Nitidulidae), longhorn beetles (Cerambycidae), leaf beetles (Chrysomelidae), rove beetles (Staphylinidae), scarabs (Scarabeidae), tumbling flower beetles (Mordellidae) and weevils (Curculionidae) are common pollen feeders of many flowers. Tumbling flower beetles are commonly found feeding on flowers of the carrot family, which purportedly contain a good pollen source. Though most beetles visit flowers solely for pollen, there are some species that desire more. Many beetle families such as checkered beetles (Cleridae; Fig. 4), and blister beetles (Meloidae) are known to use other floral resources. Indeed, some prefer to simply eat the flowers and will gnaw flower tissue, eat pollen and lick floral exudates. However, in the case of some beetles, flowers represent much more than just a place to get free food. For example, monkey beetles (Scarabaeidae), a primary pollinator of the peacock moraeas plant in South Africa, frequent flowers of this plant for pollen, nectar and mating, while beetles from South America are attracted by heat-producing Aroid plants and a series of water lilies, in which they feed and reproduce. More locally, many beetles, including soldier beetles, often use the same flower that they frequent in search of food for mating (Fig. 5). Besides using flowers as sources of food and mating, beetles sometimes use them as their residence and a place to prey on other insects. For example, predatory beetles often hide within flowers, waiting for soft-bodied insects to visit. Ladybeetles, which are well known predators, visit flowers to feast on aphids and may sip some nectar when available. Beetles may also use flower blooms as a place to hide from predators. Some beetles may also enter flowers because the temperature within their blossom is more preferable to external environmental conditions. Thus, beetle-pollinated flowers include either sufficient nutritional rewards, place to warm up, mate and hunt or serve as a refuge from natural enemies.
Fig. 4. Ornate checkered beetle Trichodes ornatus visiting a flower. Photo: J. Gallagher (CC)
Fig. 5 Soldier beetle feeding and mating on same flower. Photo: S. Nygaard (CC)
Beetle importance as pollinators
Beetles make up the largest group of pollinating animals because of their large numbers and are the most diverse group of pollinators in the US. More than 77,000 beetle species are estimated to visit flowers. Of the world’s almost 350,000 flowering plant species, beetles are believed by some to be responsible for visiting nearly 90%. Cantharophily (cross-pollination of flowers by beetles) is more common in tropical areas especially in tropical-Mediterranean regions and is often not considered important to pollination in temperate regions. Relative to this, the native paw-paws and atemoya are some of the only crops in the U.S. known to be pollinated by beetles. However, an estimated 52 native plant species are pollinated by beetles in North America north of Mexico, and there are several common temperate ornamental plants that are beetle pollinated. Beetles may also be particularly important in semi-desertic areas, such as South Africa and southern California. Beetles are important for the production of crops that are exclusively beetle-pollinated, such as atemoyas or paw-paws. To this point, a relatively recent study found that atemoya orchards in Australia directly benefit from the presence of wild beetle species in surrounding forests, and that increasing the presence of natural habitats around cropping areas improves fruit set and production. From this respect, the presence of wild beetles is an extremely valuable ecosystem service, which if present, can help some industries which rely on hand-pollination cut labor cost.
Despite contributing to the pollination of many plants, beetles’ roles in ecosystem services are generally assumed to revolve around their scavenging and organic matter decomposition. Part of this misconception may be attributable to the fact that flower-visiting beetles are less active on flowers than are many flies, butterflies and bees, and for that reason they are usually considered inefficient pollinators. Further, some scientists claim that beetle pollination is among the most destructive: most beetles eat their way through petals and other flower parts, they defecate within the flower, and then spread the mixture of feces and pollen. Still, beetles are recognized as the primary pollen transporters for numerous plant families, especially phylogenetically basal plants such as magnolias and water lilies. For this reason, it is unfair to not recognize beetles as vital pollinators who play a unique role in wild plant reproduction and food production.
Beetle floral preference
Flower description. Flowers dependent on beetle pollinators contain a foreseeable list of features that differ considerably from flowers primarily pollinated by other insect orders. Flowers visited by beetles may be large solitary (e.g. magnolias, water lilies) or clusters of small flowers (goldenrods, Fig. 6; Spirea, Fig. 7). Beetles are generally clumsy and rough fliers, compared to more delicate and/or agile flying insect pollinators (e.g. butterflies, bees, flies). To accommodate these clumsy fliers, beetle-pollinated flowers tend to be large and their rewards easily accessible. Further, the volume of individual flowers is typically large enough to accommodate several beetles within the same floral cup. Beetle-pollinated flowers usually have open corollas, solitary and heavily-constructed, though some may contain many tiny clustered flowers. Beetles also frequent flat to dish-shaped or bowl-shaped flowers, as these features provide them an easy platform for landing and often a good place for shelter.
Fig. 6. Goldenrod soldier beetle on goldenrod. Photo: D. Hill (CC)
Relatively large beetles can often damage flowers, or their pollinating parts, especially when feeding on pollen with their large mouth-parts. However, many flowers resist this damage either by producing many more flowers than what would be sufficient for reproduction, or by physically protecting the most important parts of their reproductive organs (e.g. the ovaries) from beetles. Some beetle-pollinated flowers such as some Aroids present structures that function as traps, preventing beetles from leaving the flower before the end of the plant reproductive cycle. Others floral traits that prove beneficial to beetle attraction include nocturnal (but also diurnal) blooms, plant heat production, and the presence of floral structures that provide protection to beetles.
Fig. 7. Spirea flower. Photo: S. Braun (CC)
Flower color. Although many beetles can see color and UV light, they use color mostly as a short-distance cue for floral choice. Indeed, it seems that most beetles base their long-distance floral location on floral odors. In terms of floral coloration, most beetles are attracted to dull-colored, greenish or white, and reddish brown or dark flowers. Flower size is not a good indicator of floral choice, since flowers preferred by beetles can be large or small and clustered.
Floral odors. Many flowers pollinated exclusively by beetles display strong fragrances. Odors that serve as primary beetle attractants are numerous and not always pleasant, including smells of decaying plant or animal material, fermented fruits or spices. Indeed, when locating flowers, beetles are attracted to a variety of scents and although there are not many plants pollinated primarily by beetles, flowers that do depend on them are typically characterized by the presence of discernible fragrances that acts as a primary long-distance attractant.
Pollen load and heat. Many beetle species eat pollen, so it is understandable that plants they frequent produce plenty of easily-accessible pollen. This also ensures that there is enough pollen that remains for pollinating the flower after the beetles complete their meal. In addition to pollen, beetle flowers often use heat as a reward for pollination. Some incredible plants are capable of producing heat, which attracts and likely increases the activity level of beetles while visiting their flowers. These heat-producing beetle-attracting flowers belong to families Nymphaeaceae, Illiciaceae and Magnoliaceae.
How to Protect beetles?
Planting swaths of wild flowers, native shrubs and trees, as well as urban green spaces will provide good habitats for adult beetles and other pollinators. Similarly, since some beetles deposit their eggs in soil or loose-leaf litter, it is critical to eliminate the use of synthetic fertilizers and toxic pesticides that threaten their life above and below the soil. This is especially important for soldier beetle larvae which are carnivorous, consequently foraging for aphid eggs, worms, slugs, and other prey among assorted plant debris. As they feed, soldier beetle larvae reduce the number of soft-bodied insects, such as aphids. Adopting organic land management practices such as planting pollinator habitat, conservation strips and cover crops, using mulch for weed control, and adding compost and diverse plantings to arable lands, helps to build and protect beetle biodiversity.
Summary
Though currently recognized as being among the oldest known pollinators of flowering plants, beetles usually don’t get their due as major pollinators. This is partially attributable to their suspected inability to move pollen through long distances and their destructive behavior while feeding on pollen and floral structures. Notwithstanding, beetles are central in the pollination of many plant species in temperate areas and are popularly known for their pollination services in tropical and Mediterranean ecosystems. In temperate regions, beetles contribute to the pollination of Magnolias and all spice bushes. In terms of crops, beetles are important in the production of some tropical fruits such as atemoyas and local paw-paws, allowing these crops to be produced without hand-pollination. Pollinating-beetle conservation involves protecting established ecosystems and increasing the presence of wild habitats by establishing natural resources such as wild flowers, native shrubs and trees. Financial support for the publication of this article is via USDA NIFA EIPM grant award numbers 2017-70006-27171.
Laura C. Moore^,*, Alan W. Leslie#, Cerruti RR Hooks$,* and Galen P. Dively+,* Former graduate student^, Associate Professor and Extension Specialist$, Professor Emeritus+, CMNS, Department of Entomology*, Agriculture Extension Agent, Charles County#
Introduction
Increasing floral diversity within agricultural fields has been proposed as a method to bolster natural enemies and subsequently reduce pest populations. A key factor that enhances predator and parasitoid populations is the availability of nectar and/or pollen food subsidies from flowering plants. Many natural enemies, particularly hymenopteran (wasps) parasitoids, require carbohydrates for successful reproduction and overall fitness. However, monoculture cropping systems are relatively weed-free and generally lack floral resources required by many natural enemies. A literature review showed that the successful establishment of certain parasitoids in cropping systems depended on the presence of nectar-bearing weeds. In addition to providing natural enemies nectar and pollen to eat, flowering plants can supply alternative hosts or prey, shelter, overwintering sites and a more suitable microclimate.
Many conservation projects have been implemented by farmers to increase beneficial services on arable lands. In Maryland, the opportunity to practice conservation biological control exists within the Conservation Reserve Enhancement Program (CREP). Conservation biological control is a pest management approach that manipulates agricultural systems so as to promote pest suppression by naturally occurring predators, parasitoids and pathogens. The CREP seeks to establish riparian buffers in Maryland to improve water quality, filter sediments and nutrients from runoff and provide wildlife habitat. However, these buffers can be engineered to support communities of natural enemies and serve as corridors for their movement into neighboring crops.
The aim of this study was to determine whether buffer strips could be used as insectary plants to enhance beneficial arthropods (insects and spiders) within neighboring soybeans. Insectary plants are plants grown with cash crops to attract, feed and shelter insect parasitoids and predators so as to enhance their natural control of insect pests. We monitored pest and beneficial arthropods in the buffer insectary plants and neighboring soybean plantings and tried to link arthropods found in buffer plants with pest management in neighboring soybeans.
Abbreviated Experimental Procedures
Insectary buffer test plants. Partridge pea and purple tansy are commonly used to enhance floral resources along field margins for pollinator plantings and to enhance communities of natural enemies in adjacent crops. Partridge pea is a native annual legume and is widely used in seed mixes of CREP riparian buffers because it readily reseeds itself, is competitive when grown with grass mixes, and provides nutritional seed for game birds. As an insectary plant, partridge pea has a long bloom period and each leaf petiole has an extrafloral nectary at its base, which produces nectar throughout the growing season. A diverse assemblage of pollinators and natural enemies are attracted to partridge pea. Purple tansy has a long flowering period, high-quality nectar and pollen production, and is reported as being a valuable insectary plant. Proso millet is a warm season annual grass that lacks floral resources. As such, it served as a grass control to determine how added vegetation diversity in the absence of floral resources would impact natural enemies.
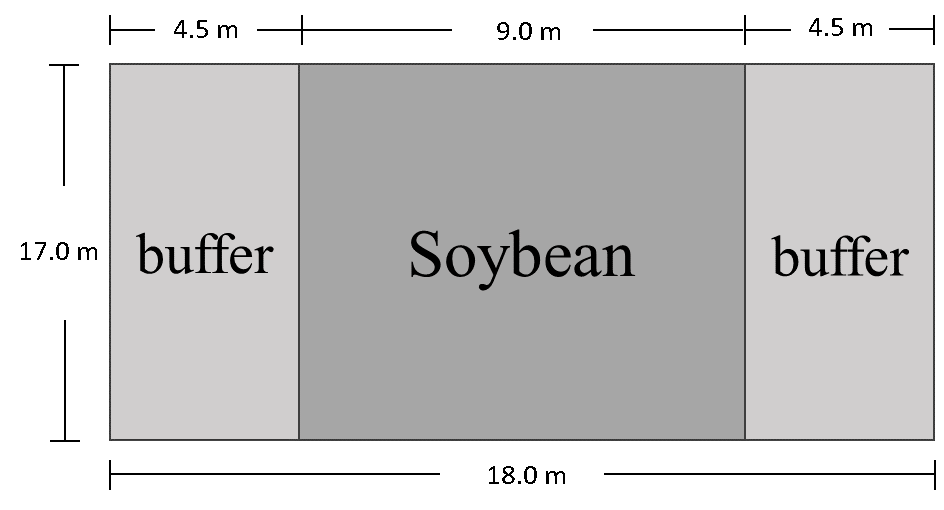
Experimental design. Field experiments were conducted over two years at the Central Maryland Research and Education Center in Beltsville, MD. In year 1, 16 plots of soybean were seeded on May 11. Each plot consisted of 20 soybean rows spaced 35 cm (15 in) apart and bordered on each side by an insectary buffer strip (Fig. 1). The test buffer strips consisted of 1) purple tansy, 2) partridge pea, 3) 50:50 seed mixture of purple tansy + partridge pea, or 4) proso millet. Each soybean plot-buffer combination was replicated four times. Seeds of partridge pea, purple tansy, and proso millet were planted with a no-till drill in rows 23 cm (9 in) apart at a rate of roughly 12,000 seeds per ha (4856 per ac) on the day soybeans were planted.
Fig. 1. Illustration of a soybean-buffer treatment plot in year 1. Soybean plots were bordered on each side with buffer insectary plants. Buffers included purple tansy, partridge pea, 50:50 seed mixture of partridge pea/purple tansy or proso millet.
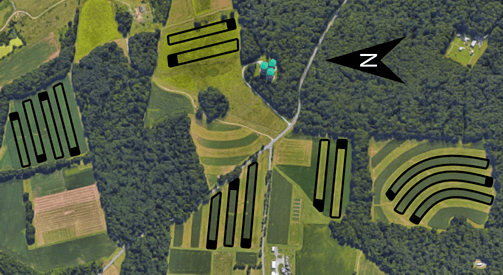
The year 1 study showed that purple tansy was unsuitable for the hot summer conditions in Maryland Thus, it was not used in the year 2 experiment, which focused solely on partridge pea as the insectary buffer plant. The year 2 experiment included 14 strip plantings of full season soybean at five different locations (Fig. 2). Soybeans were planted no-till in 75 cm (30 in) wide rows during May. Each strip was bordered at one end with a partridge pea buffer and at the other end with a mixed grass border of fescue (Festuca spp.) and orchardgrass (Dactylis spp.).
Fig. 2. Aerial view of experimental layout in year 2. Study consisted of 14 contour strips of full-season soybeans and adjoining partridge pea buffers (indicated by black polygons) at one end of each strip. Grassy areas were on opposite ends of soybean strips without a buffer.
Arthropod population assessments. Abundances of arthropods active in the plant canopy were measured with yellow sticky cards secured to bamboo poles. Further, sweep-net samples were taken in July and August to estimate green cloverworm (Hypena scabra) numbers. The green cloverworm served as a bioindicator of changes in pest populations potentially caused by enhanced natural enemy activity. The larger field size in year 2 allowed sticky cards to be placed throughout the soybean strip. One card was placed in the center of each partridge pea buffer, and additional cards were placed at distances of 3, 6, 12, 18 and 24 m (10 ft to 79 ft) from the border on both sides of each soybean strip (total of 10 sticky cards per strip). Sampling was conducted weekly or biweekly. In year 2, pitfall traps were also installed in the ground adjacent to each sticky card to estimate the abundance of surface-dwelling arthropods over 7-day intervals.
Summary of Results
Year 1 Study – comparison of four insectary buffers parasitoid abundance. Three families of parasitoids Mymaridae, Scelionidae and Trichogrammatidae comprised 83.9% of the total of parasitic wasps captured on sticky cards. Families Ceraphronidae, Braconidae and Eulophidae comprised an additional 12.5% of the wasp parasitoid group. Of these parasitoids, mymarids were the most abundant and there were 73-78% higher sticky card captures of this wasp in partridge pea compared to purple tansy and millet buffers. However, significantly fewer mymarids were captured in soybeans adjacent to partridge pea than adjacent to purple tansy or millet. Scelionid parasitoids were more abundant in millet and purple tansy buffers but their numbers were similar in soybeans regardless of the neighboring buffer type. Trichogrammatid abundance was greatest in millet early in the season and in buffers with partridge pea by season end. Two families of fly parasitoids (Tachinidae and Sarcophagidae) averaged 9.4 and 4.4 flies per sticky card in insectary buffers and soybean plots, respectively. The abundance of sarcophagid flies was significantly higher in buffers with partridge pea than millet or purple tansy alone. Similarly, soybeans adjacent to partridge pea were inhabited by more tachinids and sarcophagids than soybeans adjacent to millet or purple tansy.
Predator abundance. Overall predator abundance was significantly higher in purple tansy and millet compared to partridge pea or mixed (partridge pea + purple tansy) buffers. Mean captures per card were 5.0 in partridge pea, 6.8 in mixed, 8.5 in purple tansy, and 10.3 in millet. However. similar predator numbers were captured in soybean plots adjacent to all four buffer types.
Insect herbivores (plant feeders). Sweep net counts of green cloverworm were statistically similar in soybean plots adjacent to the four different buffer types. Overall numbers per 10 sweeps averaged 24.6, 27.0, 18.0, and 23.0 in soybeans adjacent to millet, purple tansy, mixed and partridge pea buffers, respectively. The bulk of other insect herbivores captured on sticky cards were mainly aphids, leafhoppers, planthoppers and plant bugs. Mean numbers captured per card were 86.1 (millet), 113.2 (purple tansy), 57.6 (mixed) and 53.7 (partridge pea).
3.2. Year 2 Study partridge pea vs. natural grass vegetation
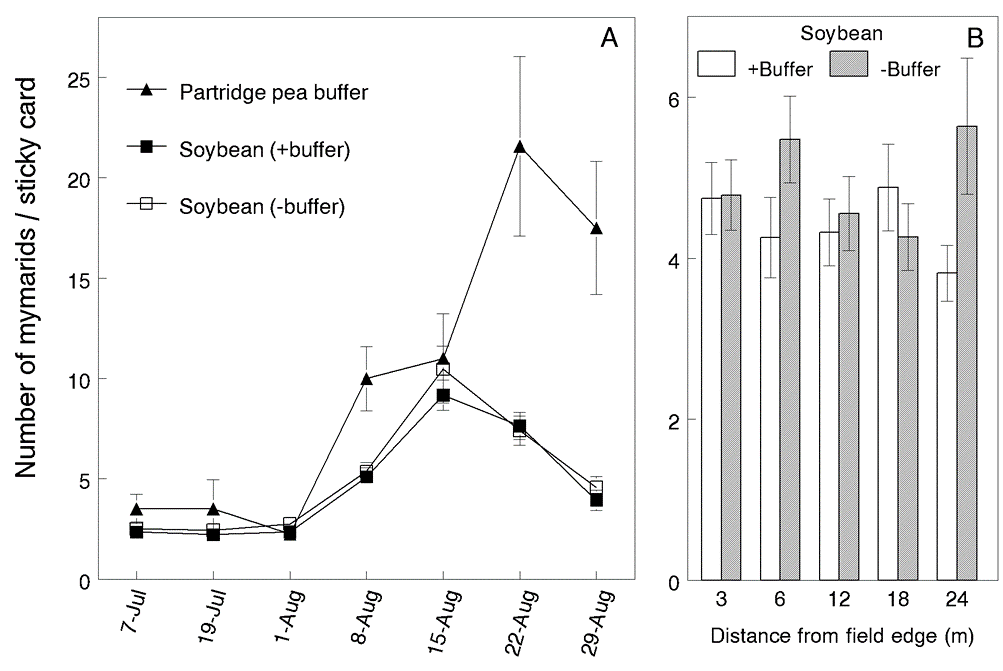
Parasitoid abundance. The most abundant parasitoids belonged to families Mymaridae, Trichogrammatidae and Scelionidae in order of abundance, and together comprised 84.3% of the total hymenopteran parasitoids captured. Each family responded differently to the partridge pea treatment. Mymarid abundance was higher overall in partridge pea buffers but did not enhance their abundance in neighboring soybeans (Fig. 3). Significantly fewer trichogrammatids were captured in partridge pea compared to numbers captured in soybean with and without the partridge pea buffer. Mean captures of dipteran parasitoids per sticky card abundance were significantly higher in soybean neighboring partridge pea, with the exception of the first and last sampling dates.
Fig. 3. A) Mean number (±SE) of mymarid parasitoids captured per sticky card in partridge pea buffer, soybean neighboring buffer, and soybean without buffer in year 2. Data for soybean were averaged over all sampling distances (3, 6, 12, 18 and 24 m) from the field edges. B) Mean number in soybean at different distances from field edges with and without a partridge pea buffer.
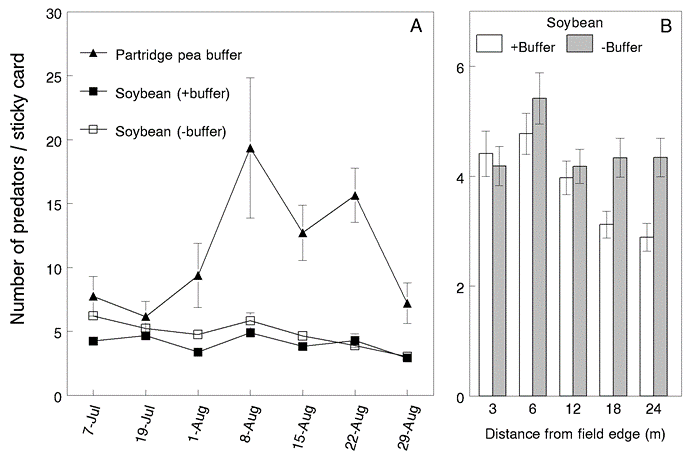
Predator abundance. Long-legged flies, minute pirate bugs, and big-eyed bugs comprised 81.4% of the total predatory arthropods captured. Soldier beetles, fireflies and lady beetles represented an additional 11.6%. Mean abundance of predators per sticky card was 11.5 ± 1.1 in buffer, 4.1 ± 0.16 in soybean neighboring buffer and 4.9 ± 0.18 in soybean without buffer. Abundance of predators was significantly lower in soybean neighboring the partridge pea buffer (Fig. 4). However, this was largely due to the activity of long-legged flies, which were more attracted to the partridge pea buffer. Still, their numbers were significantly lower in soybean strips neighboring partridge pea compared to soybeans without partridge pea buffers.
Fig. 4. A) Mean number (±SE) of arthropod predators captured per sticky card in partridge pea buffer, soybean neighboring buffer and soybean without buffer in year 2. Data for soybean were averaged over all sampling site distances (3, 6, 12, 18 and 24 m) from the field edges. B) Mean number in soybean at different distances from field edges with and without a partridge pea buffer. Arthropod predator guild consisted of long-legged flies, minute pirate bugs, big-eyed bug, soldier beetles, fireflies and lady beetles.
Insect herbivores/pests. Thrips, leafhoppers, treehoppers, froghoppers and planthoppers comprised over 95% of herbivores captured on sticky cards. The total number of herbivores per sticky card averaged 108.2 in the partridge pea buffer, 96.3 in soybean neighboring buffer, and 96.4 in soybean without buffer. Thus, herbivore numbers did not differ significantly in the buffer and soybeans.
Pitfall trap predators. A total of 56,296 arthropods were identified from pitfall trap samples. Of predators captured in pitfall traps, ants, spiders, soldier beetle larvae, rove beetle adults and larvae, and ground beetle adults and larvae were the most abundant. Ant numbers were significantly lower in soybeans neighboring partridge pea on all sampling dates.
Discussion
Year 1 study was conducted to determine if pure and mixed buffer strips of partridge pea and purple tansy could attract greater numbers of beneficial arthropods than non-floral strips of millet, and whether these buffers enhance beneficial arthropod abundance in neighboring soybeans. Purple tansy was not a suitable insectary plant as it was not well adapted to the seasonal period of the study in Maryland. Furthermore, purple tansy would probably be less desirable to establish and maintain as a buffer strip due to its relatively high seed price, slow growth characteristic and greater susceptibility to weed competition. Moreover, purple tansy was quickly out-competed by partridge pea in the mixed planting to the extent that the pure and mixed buffers containing partridge pea attracted similar arthropod communities.
Overall, results consistently showed that partridge pea attracted and supported high populations of natural enemies and potential hosts and prey, with abundances significantly greater than levels found in adjacent soybeans. Sticky card captures of wasp and fly parasitoids in year 1 were more than 70% higher overall in buffers containing partridge pea compared to other buffer types. Similarly, populations of all beneficial arthropods captured by sticky card and pitfall sampling in year 2 were approximately 80 to 72% higher, respectively, in partridge pea buffers compared to the soybean crop.
Parasitoids. Mymarid wasps were notably the most common parasitoids captured on sticky cards and consistently more abundant in partridge pea compared to soybean. These tiny wasps parasitize insect eggs in concealed sites within plant tissues or the soil and are important natural control agents of economically important leafhopper pests. In year 1, mymarids reached levels in partridge pea buffers that were four-fold higher than those in soybean plots, yet significantly lower levels of mymarids were captured in soybean adjoining these buffers. This suggests that the partridge pea lured mymarids from neighboring soybeans. High numbers of mymarids were also captured in partridge pea in year 2 but their abundance in soybeans was not enhanced. This suggests that partridge pea may provide some parasitoids and their associated hosts with all resources required for survival and reproduction. This would in effect provide no incentive for these parasitoids to forage within neighboring crops.
Most fly parasitoids found on sticky cards were tachinids or sarcophagids. The vast majority of hosts of tachinid flies are plant-feeding insects. Their level of parasitism can vary greatly, from less than 1% to approaching 100%, depending on such factors as the size of a host and parasitoid population, and environmental conditions. During both study years, their overall abundance in partridge pea was 62.3% higher than levels in soybean. In year 2, this effect was heightened at the field edge next to buffers, suggesting that higher numbers of parasitic flies encroached into the neighboring soybeans but enter only a short distance within the crop.
Predators. In year 1, predators captured on sticky cards were 65% more abundant in the millet and purple tansy buffers. This response was mainly attributed to the abundance of long-legged flies. These predatory flies hover while searching for small, soft-bodied arthropods, particularly other flies, aphids, spider mites, larvae of small insects and thrips. However, abundances of long-legged flies in soybean plots were not affected by buffer type in year 1. Long-legged flies were also the predominant predators active in the plant canopy in year 2, with overall numbers 2-3 times higher in partridge pea buffers compared to levels found in soybeans. However, their abundance was significantly lower in soybean neighboring partridge pea, particularly at sampling sites closest to the field edge. This is further evidence that the partridge pea acted as a natural enemy sink.
Of the ground-dwelling predators captured by pitfall traps, ants were the predominant group and their abundance was significantly higher in partridge pea than adjoining soybeans. Their numbers were significantly lower in soybean plantings adjacent to partridge pea than grassy check treatment on all sampling dates, implying again that partridge pea acted as a natural enemy sink by luring ants away from soybean. Populations of other ground-dwelling predators, which consisted mainly of spiders, rove beetles, soldier beetles and ground beetles, showed a definite preference for partridge pea compared to soybeans. However, their abundances in the crop were not affected by partridge pea presence.
Herbivores. Sticky card captures each study year indicated that partridge pea harbored significantly more insect herbivores compared to soybean. The majority of herbivores were aphids, leafhoppers, planthoppers and plant bugs. In year 1, number of green cloverworm, as well as other herbivores in soybean were similar regardless of the buffer treatment.
Conclusion
This study demonstrated that partridge pea provides floral resources and alternative food for a diverse community of natural enemies and herbivores. However, its presence as a monoculture buffer did not result in increased number of major natural enemies in neighboring soybeans. Taken together, partridge pea planted as a monoculture acted more as a natural enemy sink by attracting beneficial arthropods away from soybean, potentially decreasing natural control efforts. For this reason, a monoculture of partridge pea may not be an ideal insectary planting if the ultimate goal is to maximize natural enemy efficacy in neighboring soybean fields.
In conservation reserve practices, monocultures of partridge pea are more commonly planted as a wildlife habitat to provide food for bobwhite quail and other wildlife and as flowering habitat for different pollinator taxa. Because the foliage is potentially poisonous to cattle and re-seeding plants can aggressively fill in voids when used as part of a seed mix, conservationists recommend for herbaceous riparian buffers that the total seed mix consist of no less than 1% and no more than 4% partridge pea. However, decisions about the deployment of insectary plants as a monoculture or part of a riparian buffer mix planting should take into consideration the attractiveness and resources provided to natural enemies and their hosts/prey by the insectary habitat in comparison to those provided by the neighboring cash crop. Simple addition of a highly attractive flowering buffer adjacent to a crop could be counterintuitive to natural biological control efforts.
Acknowledgements
Financial support for field studies and publishing results was provided by the Northeast Sustainable Agriculture Research and Education Grants Program, Maryland Soybean Board and USDA NIFA EIPM grant number 2017-70006-27171.
Maria Cramer, Edwin Afful, Galen Dively, and Kelly Hamby Department of Entomology, University of Maryland
Overview
Background: Due to their low cost, pyrethroid insecticides are often applied when other chemical applications are made. For example, they may be included in tank mixes with herbicides in early whorl corn and with fungicides during tasseling. These pyrethroid sprays often target stink bugs; however, the timing of these treatments is not ideal for stink bug management. Pyrethoid insecticides may harm beneficial insects that help keep pest populations in check and repeated use of pyrethroids can contribute to insecticide resistance.
Methods: In this study, we examined the effect of Bifenture EC® (pyrethroid active ingredient: bifenthrin) applied with herbicides in V6 corn and with fungicides in tasseling corn. We evaluated impacts on pests and beneficials at both application timings. Yield was measured at harvest.
Preliminary Results:At both application timings, Bifenture EC® did not improve insect pest management because pests were not present at economic levels. We did not find evidence for flare-ups of aphids or spider mites, but a rainy late summer made it unlikely that we would see many of these pests. There were no yield differences between the treatments.
Background
As a result of the low cost of pyrethroid insecticides, preventative applications are common, especially in tank mixes with other routine chemical inputs, such as herbicides and fungicides. However, lower grain prices and low insect pest pressure make it less likely that pyrethroid applications will provide economic returns. Bt hybrids1 and neonicotinoid seed treatments control many of the pests targeted by pyrethroid insecticides. Because they have broad spectrum activity, pyrethroids can negatively impact natural enemies2 which can result in flare-ups of secondary pests3. Tank mix timings may be less effective than applying when insect populations reach threshold. For example, when pyrethroids are combined with herbicide applications, they are too late to control early-season stink bugs and other seedling pests. When pyrethroids are combined with fungicide sprays at tasseling, few insect pests are present at damaging levels. Stink bugs may feed on the developing ear at this time, causing deformed “cowhorned” ears; however, this is rarely a problem in Maryland and stink bug damage is generally not economic throughout a field because feeding is primarily concentrated at the field edge4. Insecticide applications at tasseling have a high potential to affect beneficial insects, especially pollinators and natural enemies that are attracted to corn pollen.
Objectives: Our objectives were to determine the effect of pyrethroids applied preventatively in tank-mixes on corn pests, beneficials, and yield.
Methods: This study was conducted in 2018 and 2019 at the University of Maryland research farm in Beltsville, MD. For each application timing, we planted four replicate plots of a standard Bt field corn hybrid, DeKalb 55-84 RIB (SmartStax RIB complete Bt insect control in addition to fungicide and insecticide seed treatments) at 29,999 seeds per acre. Standard agronomic practices for the region were used.
Herbicide (same as above) + Insecticide (Bifenture EC® 6.4 oz/acre)
Treatments were applied at V6/V7. We visually surveyed corn plants for pest and beneficial insects before and after application. We also placed sentinel European corn borer (ECB) egg masses in the field to assess predation rates before and after treatment.
The fungicide timing compared two treatments:
Fungicide alone (Trivapro® 13.7 oz/acre)
Fungicide (same as above) + Insecticide (Bifenture EC® 6.4 oz/acre)
Treatments were applied at green silk. We inspected the ear zone and silks for pests and beneficial insects before application. After application, we recorded the number of ears with pest damage and the kernel area damaged. We also counted stink bug adults and cowhorned ears. Six weeks after application, we visually assessed plants for spider mite and aphid colonies.
Sampling for pests and beneficials (left) and; sentinel European corn borer egg mass (right).
Results
In the herbicide-timing study in 2019 we observed no effect on beneficial insects from the treatments (Figure 1). The most abundant beneficial species were minute pirate bugs and pink spotted lady beetles, which are very mobile and may have recolonized treated plots after treatment. Similarly, treatments did not affect predation on the sentinel egg masses, suggesting that the pyrethroid application may not have affected predators’ ability to locate and consume eggs. Across the treatments, 30-50% of egg masses were consumed by predators.
Minute pirate bug on European corn borer egg mass.
The treatments did not impact the number of beneficials at the herbicide timing (N.S.). The pyrethroid insecticide significantly reduced the number of plant hoppers and plant bugs from less than 4 per plant on average to less than 2 per plant (significantly different p<0.05, *), though these insects are not economic pests at this stage. There were never more than 2 stink bugs per 90 plants, well below the treatment threshold of 13 per 100 plants4.
In the fungicide-timing study in 2019, beneficials, especially minute pirate bugs, were abundant at the time of application (3 in every 10 plants), while stink bugs, the presumed target pest, were very rare (1 stink bug in every 68 plants). In 2018, stink bugs were similarly scarce. Overall pest abundance was low (1 in every 35 plants). After application, there was no difference in the incidence or amount of the corn ear damaged by worms, stink bugs, or sap beetles between treatments. Average stink bug and earworm incidence was roughly 1 in 10 ears, while sap beetle was even less frequent. Cowhorned ears and adult stink bugs were almost non-existent in both treatments.
Six weeks after application we found no differences in aphid or spider mite populations between the treatments, suggesting that pyrethroid applications at tasseling did not cause secondary pest outbreaks. We sampled after a period of dry weather; however, the late summer was rainy at Beltsville, which likely suppressed spider mite and aphid populations. Under drought-stress, reductions in the natural enemy population from pyrethroid use might contribute to flare-ups of aphids and spider mites.
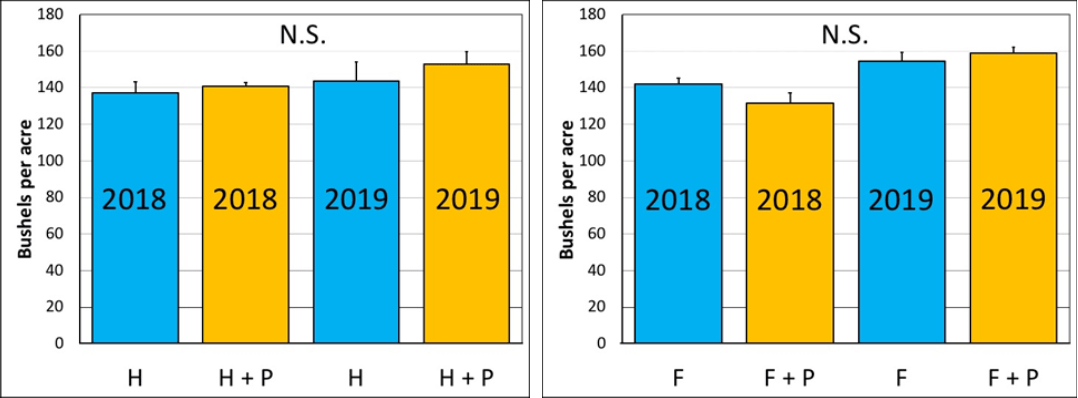
Figure 1. Herbicide timing. July 3, 2019, Beltsville MD. Mean number of insects per 10 plants in V7 corn after treatment. N.S.=not significant. H=herbicide; P=pyrethroid.
Yield
For the herbicide timing and fungicide-timing (Figure 2) studies, treatments did not affect yields in either 2018 or 2019.
Conclusions
Figure 2. Herbicide timing (left) and fungicide timing (right), 2018 and 2019, Beltsville MD. Mean yield per acre under two treatments. Yields were not significantly different by treatment in either study. For the fungicide-timing study, 2019 yields were significantly higher than in 2018. N.S.=Not Significant. H=Herbicide; F=Fungicide; P=Pyrethroid.
Results from the 2018 and 2019 studies suggest that pyrethroid applications do not provide yield benefits in corn when tank-mixed with herbicides or fungicides, likely due to the lack of insect pest pressure at these spray timings. Beneficial insects were abundant in the crop at each of these timings and did not appear to be affected by the pyrethroids in the herbicide plots. Repeated preventative use of pyrethroids in the same field could potentially hinder the natural biocontrol of corn pests.
Lady beetle larva (a predatory insect) in silks.
Sources
1 DiFonzo, C. 2017. Handy Bt Trait Table for U.S. Corn Production, http://msuent.com/assets/pdf/BtTraitTable15March2017.pdf
2Croft, B.A., M.E. Whalon. 1982. Selective toxicity of pyrethroid insecticides to arthropod natural enemies and pests of agricultural crops. Entomophaga. 27(1): 3-21.
3Reisig, D.C., J.S. Bacheler, D.A. Herbert, T. Kuhar, S. Malone, C. Philips, R. Weisz. 2012.Efficacy and value of prophylactic vs. integrated pest management approaches for management of cereal leaf beetle (Coleoptera: Chrysomelidae) in wheat and ramifications for adoption by growers. J. Econ. Entomol. 105(5): 1612-1619
4Reisig, D.C. 2018. New stink bug thresholds in corn, https://entomology.ces.ncsu.edu/2018/04/new-stink-bug-thresholds-in-corn/
By Emily Zobel, Agriculture Agent Associate University of Maryland Extension, Dorchester County
Grass sawflies can cause damage to both barley and wheat. Grass sawfly larvae hatch and feed on the lower leaf blade in early May. As it grows, the larvae will climb and continue to feed on stems and leaves. Stem and head clipping often occur before leaf feeding is complete and/or the grain reaches physiological maturity. A sawfly larva may clip >10 heads before it matures and drops to the ground.
The grass sawfly larvae can be distinguished from other pests by their solid green color, amber head with a brown band and many legs. Sawfly larvae are often mistaken as caterpillars, but are actually in the insect order Hymenoptera (wasp, bees, and ants). You can tell the difference between a caterpillar and sawfly larvae by counting the number of prologs. Caterpillars will have 5 or fewer pairs of pro-legs while sawfly larvae will have 6 or more pairs of pro-legs (Figure 1).
Figure 1. Proleg comparison between a sawfly larva and an armyworm. Sawflies have 6 or more pairs of prolegs. Images: University of Maryland and University of Delaware.
You can scout for sawfly larvae and armyworms by shaking stems. Shake both sides of 5 linear feet of row and examine any worms that fall off between the 2 rows and note any head clipping. Repeat for at least 10 sites. The threshold for sawfly larva and armyworm is when the larvae number more than 0.4 per linear ft. of row or 0.7 per square ft. and are smaller than ¾ inch. If the larvae are over 1 inch, have a dark bar on the head capsule, and have clipped many heads, it is probably too late to treat.
Some chemical control options are as follows: BaythroidXL (beta-cyfluthrin), Besiege (lambda-cyhalothrin+ chlorantraniliprole), Blackhawk 36WG(spinosad), Mustang MAXX 0.8 EC (zeta-cypermethrin), Radiant SC (spinetoram, Warrior II (lambda-cyhalothrin), and Tombstone 2 EC (cyfluthrin). Remember the label is the law. Be sure to read the label before making any pesticide applications and observe all label restrictions. Please check the label for rates and pre-harvest interval and restrictions.
Article adapted from information from University of Maryland and University of Delaware.
Bill Cissel, Extension Agent – Integrated Pest Management, University of Delaware; bcissel@udel.edu
Congratulations to John Comegys for correctly identifying the insect as a minute pirate bug or insidious flower bug and for being selected to be entered into the end of season raffle for $100 not once but five times. Everyone else who guessed correctly will also have their name entered into the raffle. Click on the Guess the Pest logo to participate in this week’s Guess the Pest challenge!
All bugs suck, but not all bugs are bad.
The minute pirate bug or insidious flower bug may look like a beetle but it is actually a “bug” meaning it has piercing-sucking mouth parts. This insect is usually considered a beneficial insect and not a pest but has been known to bite humans, inflicting a sharp and painful bite. It is commonly observed in greenhouses preying on thrips, various insect eggs, and other small, soft bodied arthropods.
The larger than life photo is also misleading and the adult is actually small, only about 1/16” in length. The immature bugs called nymphs resemble the adults in shape but lack wings and are a reddish – orange color. The nymphs, similar to the adults are also predators and feed on small arthropods.